
ARTIGOS
Como é o ciclo do vida do HIV?

Por Dr. Richard Portier
em 10 de abril de 2026.
No terceiro capítulo do livro “Fundamentals of HIV Medicine 2025 Edition”, os autores abordam o ciclo de multiplicação do HIV.
No HIV maduro, o capsídeo viral contém duas moléculas de RNA de fita simples e as enzimas necessárias à infecção: transcriptase reversa, integrase e protease, além das proteínas acessórias. O capsídeo é envolvido por uma proteína estrutural denominada matriz (p17). Esta é envolvida pelo envelope viral, composto por uma bicamada fosfolipídica derivada da membrana da célula hospedeira, que contém as glicoproteínas virais gp120 e gp41. As glicoproteínas gp120 possuem um alto nível de variabilidade, o que permite ao vírus escapar de anticorpos neutralizantes produzidos pelo sistema imunológico.
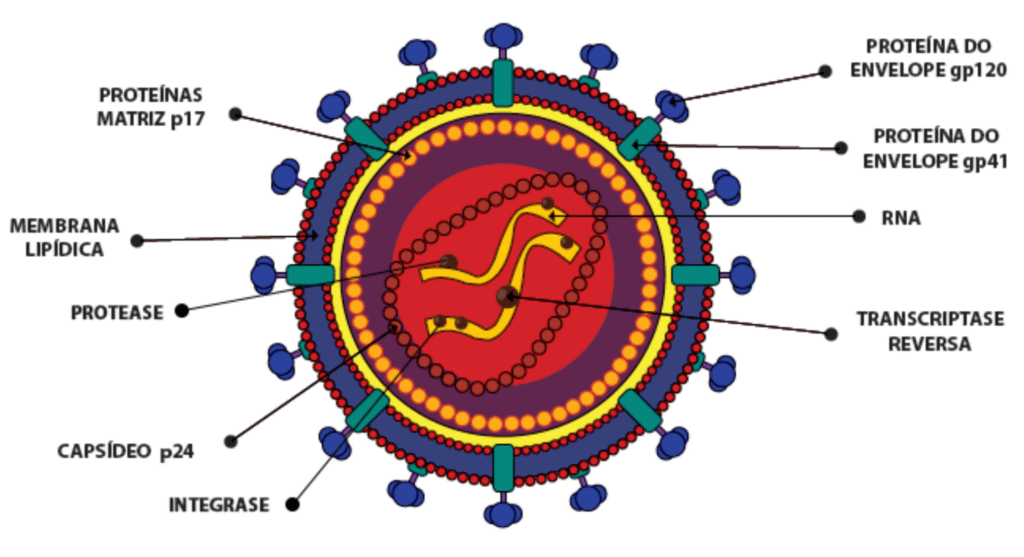
Imagem retirada do protocolo “Diagnóstico do HIV”do Ministério da saúde, 2014.
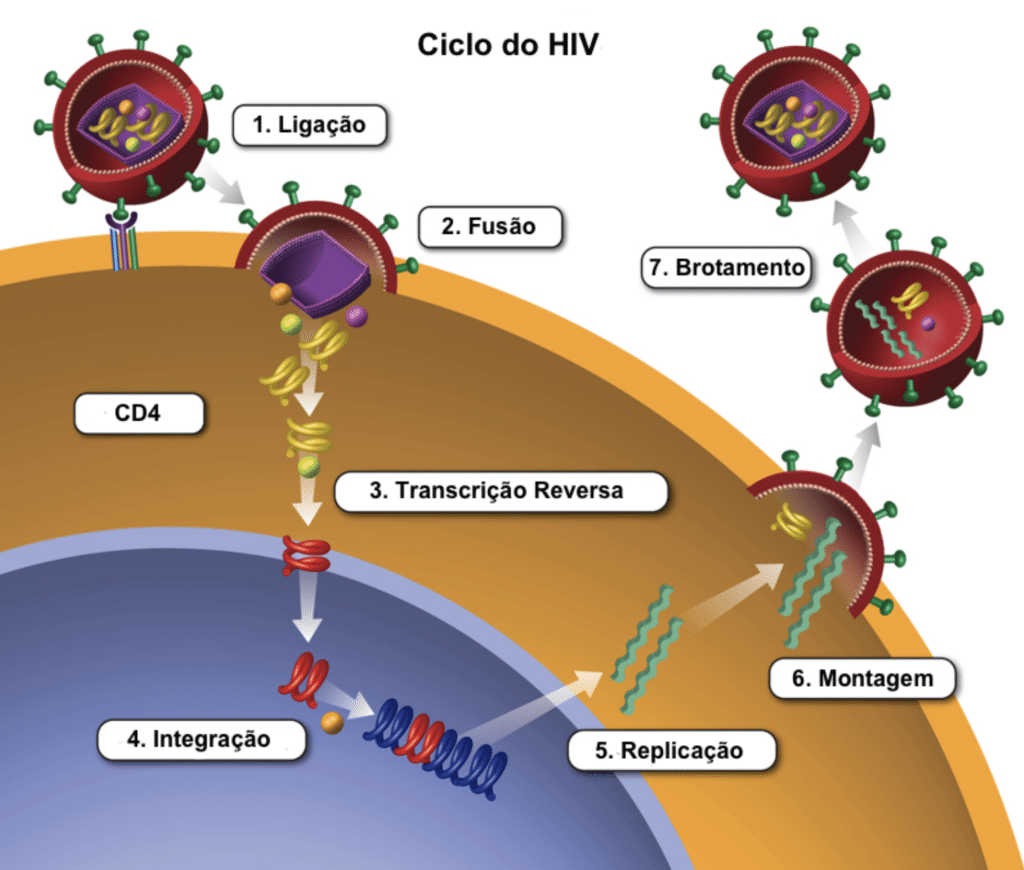
O Ciclo de Replicação Viral
Entrada Viral e Fusão
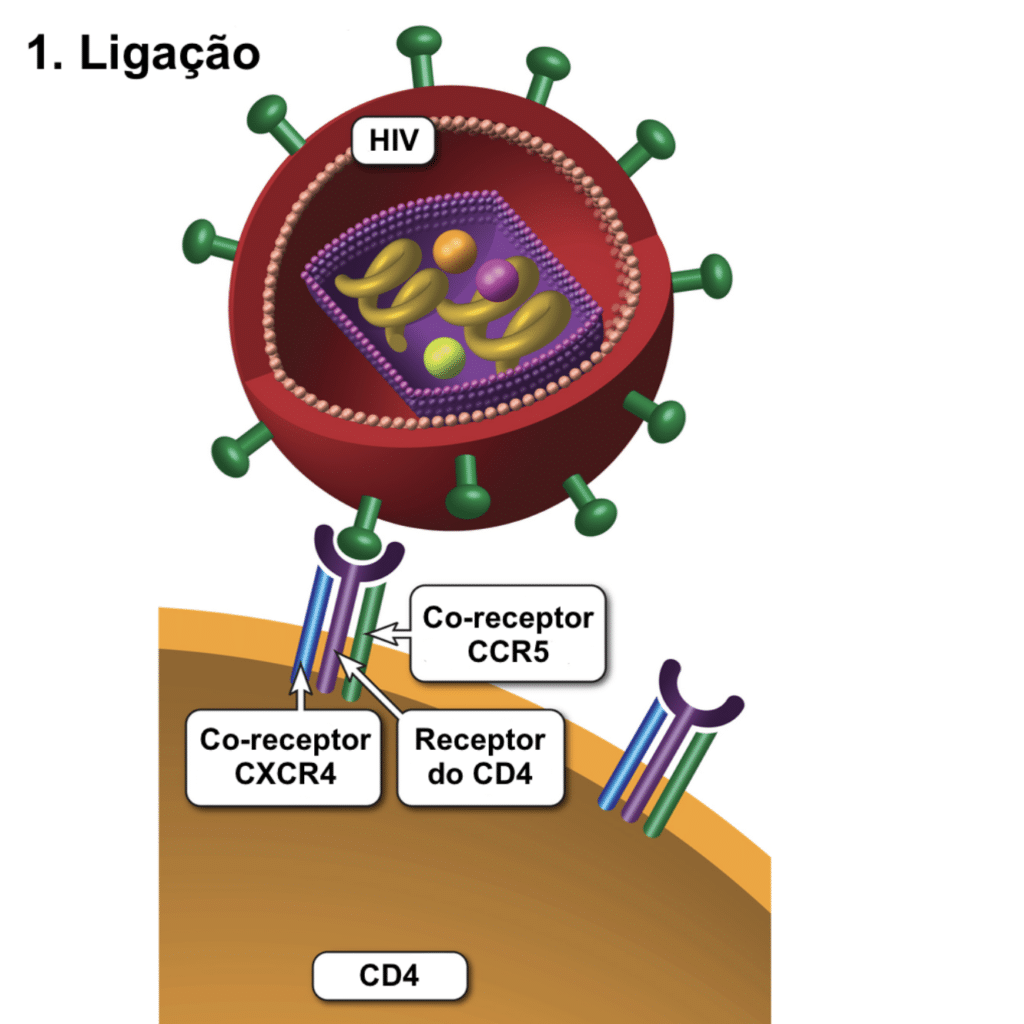
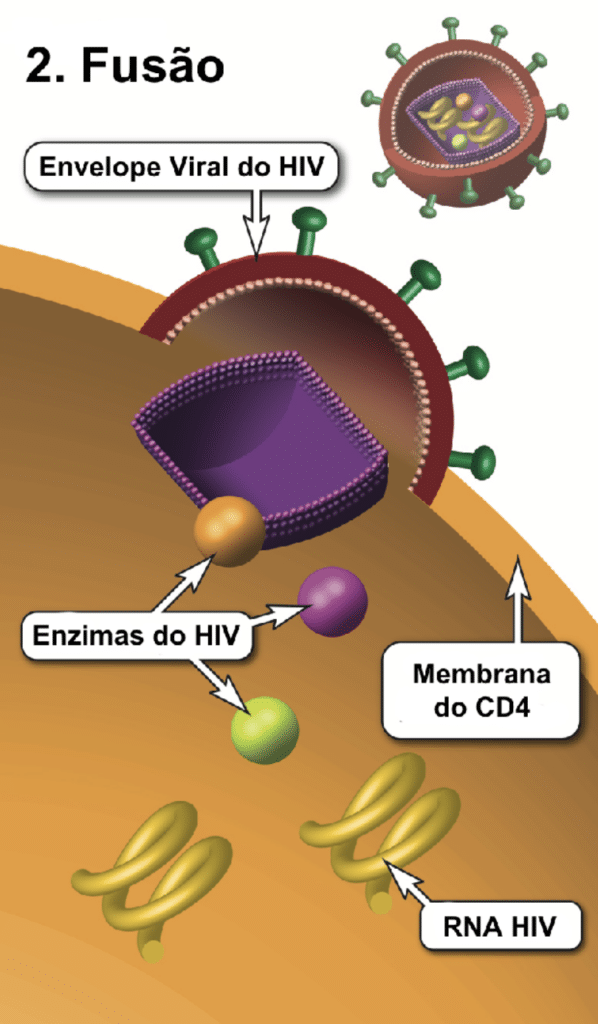
O envelope viral contém as proteínas para a fusão celular, iniciando a infecção do linfócito T CD4+. A gp120 se liga com alta afinidade ao receptor CD4. Esta ligação induz uma mudança conformacional, expondo seus sítios de ligação para os correceptores CCR5 ou CXCR4 na superfície celular. A ligação ao correceptor permite que a gp41 insira seu peptídeo hidrofóbico na membrana, formando um poro por onde o capsídeo entra. A gp41 é o alvo dos antirretrovirais (ARV) inibidores de fusão.
O uso dos coreceptores varia de acordo com a cepa viral. Cepas que usam CCR5 ou CXCR4 são classificadas como R5-trópicas ou X4-trópicas, respectivamente. Pessoas com a mutação genética CCR5Δ32 não expressam esse correceptor e são altamente resistentes ao HIV R5-trópico. Antirretrovirais antagonistas de CCR5 só funcionam para cepas R5-trópicas. Houve raros casos de remissão (cura funcional) após transplante de medula de doadores homozigotos CCR5Δ32.
Transcrição Reversa e Integração
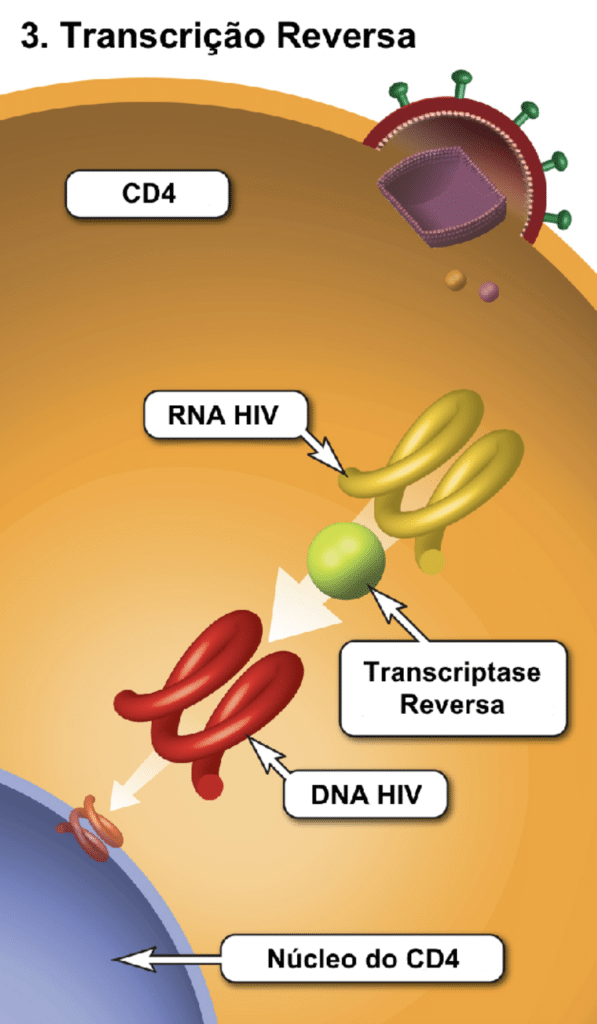
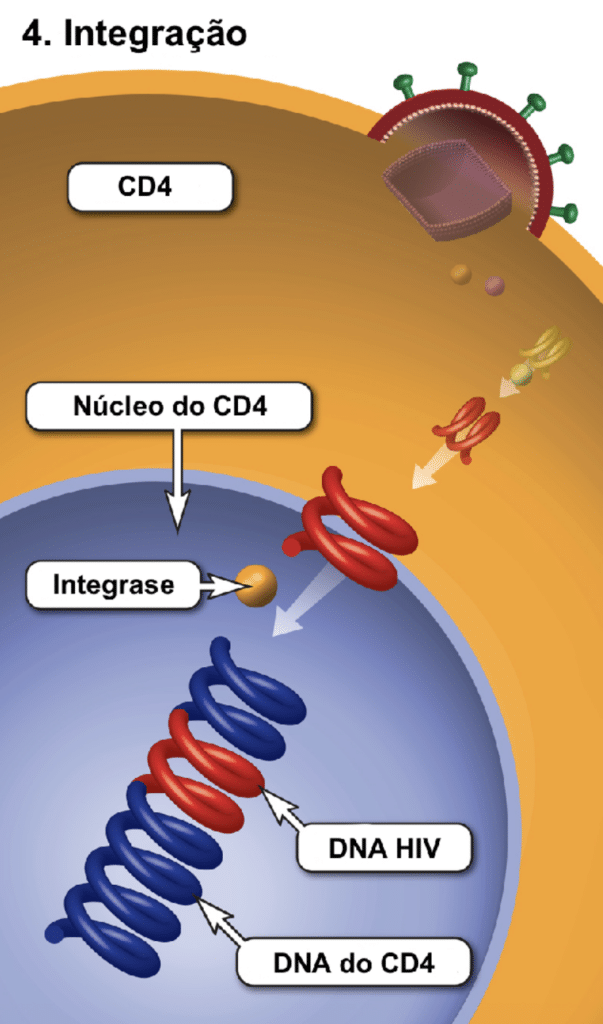
Após a fusão, ocorre o desencapsidamento. A enzima transcriptase reversa transforma o RNA viral em DNA viral. O sistema imune inato tenta bloquear essa etapa produzindo a enzima APOBEC3G, mas a proteína viral Vif a neutraliza e a degrada. O DNA viral recém-sintetizado é então transportado ao núcleo e integrado ao DNA da célula pela enzima integrase. Neste estágio, ele passa a ser chamado de provírus.
Produção Viral
Uma vez integrado, o provírus pode permanecer latente ou sofrer expressão ativa. Na expressão ativa, o DNA viral é transcrito em mRNA, que é processado e traduzido em proteínas virais progenitoras. Em um ciclo de feedback positivo, a proteína regulatória Tat promove ainda mais a transcrição viral. A proteína estrutural Gag promove a montagem dessas proteínas progenitoras para formar novos vírions.
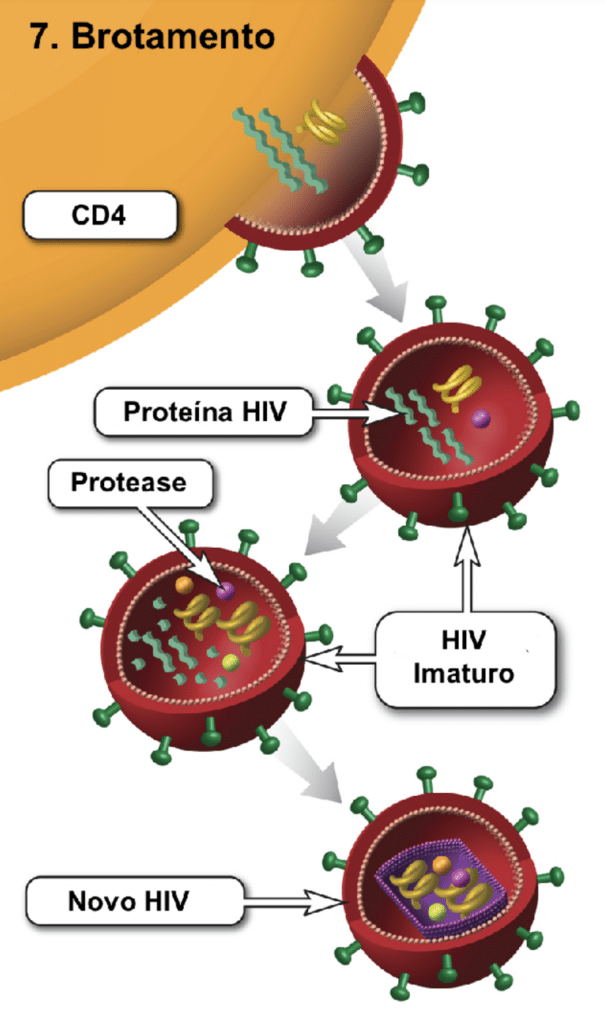
Brotamento e Maturação
A montagem ocorre na membrana celular, e as partículas deixam a célula por brotamento, incorporando partes da membrana celular ricas em colesterol e esfingolipídios. Durante ou logo após o brotamento, a enzima protease atua amadurecendo o vírion, tornando-o infeccioso. O tempo de geração do HIV-1 in vivo é de 2,5 dias, e a meia-vida dos linfócitos T CD4+ infectados é de apenas 0,7 dias.
Estabelecimento da Infecção
Na transmissão sexual, o HIV atravessa a barreira epitelial por rupturas físicas causadas por trauma ou outras infecções sexualmente transmissíveis. O vírus também pode atravessar mucosas intactas via células dendríticas (especialmente as células de Langerhans) ou por transcitose no trato gastrointestinal.
Ao cruzar a barreira epitelial, o vírus encontra múltiplas células-alvo. A infecção inicial é frequentemente propagada pelas células dendríticas, que entregam o HIV aos linfócitos T CD4+. Alternativamente, os vírions podem infectar diretamente células CD4+ locais e células derivadas de monócitos.
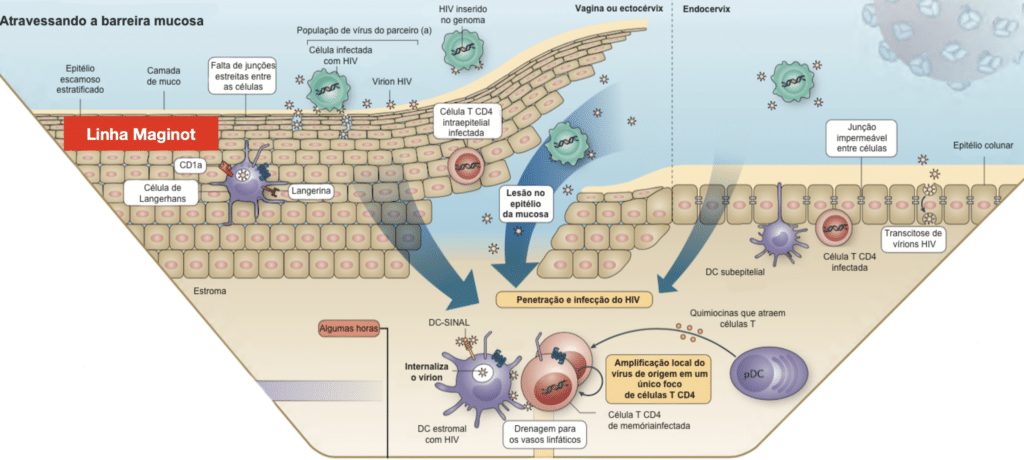
Imagem adaptada de The immune response to HIV. Nina Bhardwaj, Florian Hladik and Susan Moir. 2012.
A fase eclipse é o período em que o vírus permanece indetectável no sangue (aproximadamente 10 dias). Uma vez estabelecido no ponto de entrada, ele se expande rapidamente para os linfonodos regionais e, através da corrente sanguínea, dissemina-se pelo corpo. Através de uma reação em cadeia chamada “sinapse virológica”, células dendríticas infectadas aumentam o recrutamento de mais linfócitos suscetíveis, espalhando a infecção
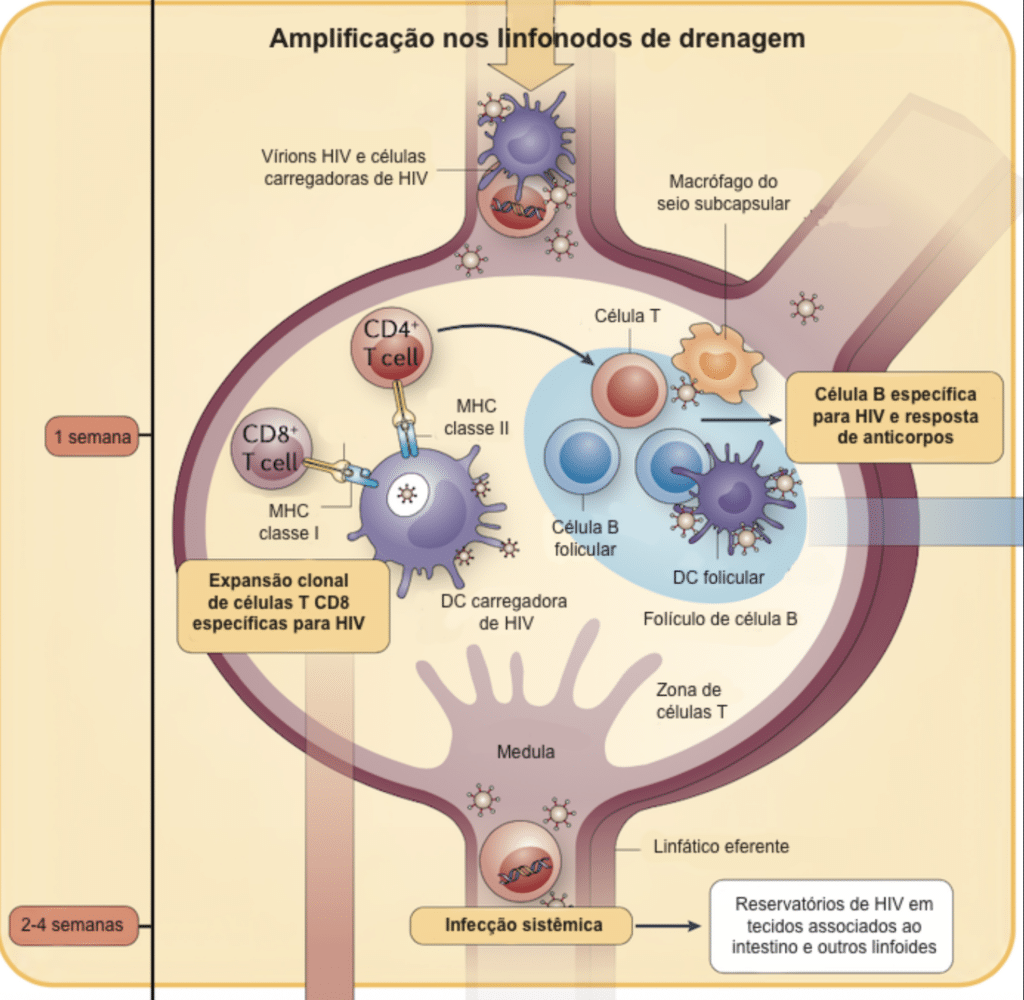
Imagem adaptada de The immune response to HIV. Nina Bhardwaj, Florian Hladik and Susan Moir. 2012.
Infecção Aguda e Reservatórios
Com a viremia alta, a contagem de linfócitos T CD4+ no sangue cai drasticamente devido a:
- Destruição direta das células pelo HIV;
- Aumento da apoptose celular;
- Destruição por linfócitos T CD8+;
- Produção reduzida na medula óssea;
- Redistribuição de células infectadas para o tecido linfoide.
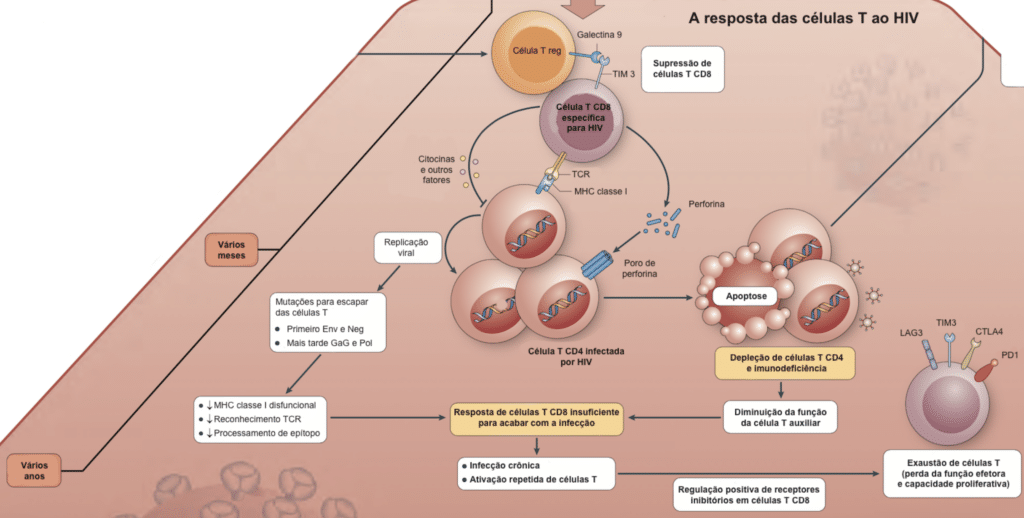
Imagem adaptada de The immune response to HIV. Nina Bhardwaj, Florian Hladik and Susan Moir. 2012.
As células NK (Natural Killers) desempenham um papel importante na infecção aguda, estimulando a destruição de células infectadas e secretando citocinas para limitar a infecção.
A ativação crônica e a replicação causam fibrose na arquitetura linfoide, levando a uma recuperação incompleta da imunidade mesmo após o início do tratamento. É aqui que o reservatório de células latentemente infectadas é estabelecido (principalmente no intestino, tecidos linfoides periféricos, tecido adiposo e micróglia cerebral). Mesmo diagnosticado na fase aguda, o tratamento imediato pode atenuar, mas não reverter totalmente, esses reservatórios, que são a principal barreira para a cura. Como esses provírus inativos não replicam, não utilizam as enzimas que os medicamentos antirretrovirais atacam.
Latência Viral Após o Início do Tratamento
Ao iniciar a terapia antirretroviral (TARV), a queda da carga viral ocorre em quatro fases distintas e contínuas. A primeira fase concentra-se nos 10 dias iniciais, apresentando uma queda rápida decorrente do bloqueio da replicação e da morte dos linfócitos T CD4+ infectados, que possuem uma meia-vida curta de apenas 1 a 2 dias. A segunda fase estende-se de semanas a meses e é marcada por um declínio mais lento, correspondendo à eliminação de macrófagos infectados e linfócitos de vida mais longa.
Na terceira fase, que pode durar de meses a anos, a carga viral continua a declinar de maneira muito lenta, o que provavelmente representa a morte natural e gradual de linfócitos latentemente infectados. Por fim, a quarta e última fase é de estabilização. Ela ocorre entre 4 e 5 anos após o início da terapia e reflete o equilíbrio entre a ativação (ou produção residual) do vírus e a sua eliminação constante pelo organismo e pelos medicamentos.
Apesar da carga viral indetectável no sangue, a proporção de linfócitos T CD4+ de longa vida que abrigam o provírus inativo declina de forma extremamente lenta, com uma meia-vida estimada de 44 meses. Por essa estimativa matemática, a erradicação total do HIV exigiria mais de 70 anos de terapia ininterrupta. Se a medicação for suspensa, essas células atuam como um reservatório a partir do qual o vírus pode reativar
Diversidade Viral
Durante a infecção não tratada, o HIV se replica a uma taxa extraordinária: cerca de 10 bilhões de novas partículas virais por dia. Como a enzima transcriptase reversa carece de um mecanismo de revisão de erros, ocorrem mutações constantes no genoma, alterando proteínas e funções virais. Outro mecanismo de diversidade é a recombinação viral, que ocorre quando uma célula é coinfectada com duas cepas diferentes do vírus, misturando seus materiais genéticos.
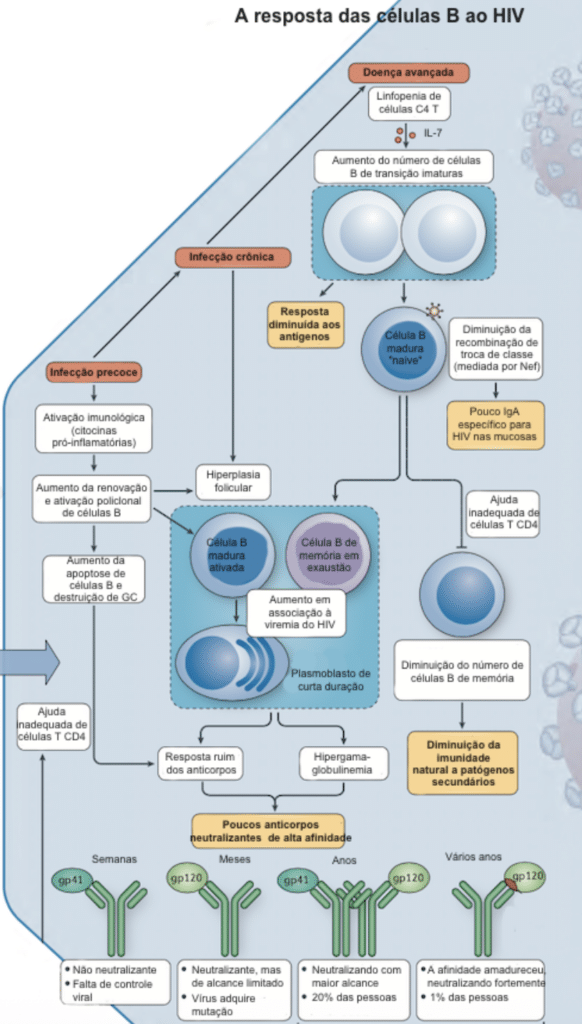
Imagem adaptada de The immune response to HIV. Nina Bhardwaj, Florian Hladik and Susan Moir. 2012.
A diversidade genética observada em um único indivíduo após cinco anos de infecção sem tratamento é aproximadamente equivalente à observada no vírus Influenza A em todo o mundo durante um ano. É essa impressionante capacidade de mutação e diversificação (que, curiosamente, é muito maior durante a infecção aguda do que na crônica) que tem impedido a descoberta de uma vacina altamente eficaz contra o HIV até os dias de hoje.





Nenhuma informação desta página substitui a consulta médica. Nunca altere seu tratamento sem antes consultar um médico ou profissional de saúde. Apenas esse profissional poderá avaliar detalhadamente sua situação clínica e decidir se você está apto a essas mudanças. Portanto, para a sua segurança, o acompanhamento médico é imprescindível.
© 2026 Richard Portier. CNPJ 31.671.711/0001-11. Todos os direitos reservados. Termos de uso.